
在大多数脊椎动物的大脑中五羟色胺能胺神经元都位于中缝核(Raphe nuclei)中。
位于中缝核中的五羟色胺能胺神经元受到来自大脑中许多区域的投射,主要包括来自腹内侧前额叶皮质(Ventromedial prefrontal cortex, vmPFC)和扣带回(Cingulate cortex)的谷氨酸能纤维和GABA能纤维,来自下丘脑视前区(Preoptic area)和延髓头端腹外侧核团(Rostral ventromedial medulla)的GABA能纤维。
不同类型的五羟色胺受体广泛地位于脑中不同类型的神经元上。
其中最重要的受体是:5-HT1A受体,5-HT2A受体和5-HT2C受体。
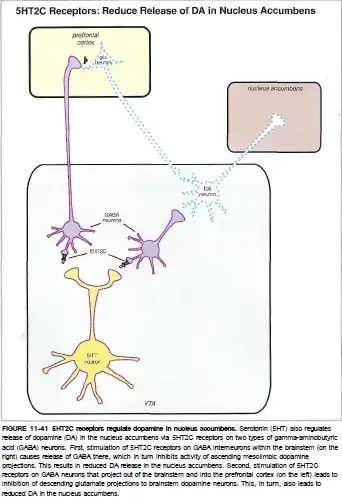
5-HT1A受体主要位于五羟色胺能神经元本身的突触膜上,额叶/顶叶皮层和海马的谷氨酸神经元上,纹状体和中脑中的多巴胺能/去甲肾上腺素能/乙酰胆碱能神经元上。
当它被激活时,可以抑制血清素本身的释放,抑制谷氨酸能锥体神经元,也可以增加多巴胺/去甲肾上腺素/乙酰胆碱的释放。
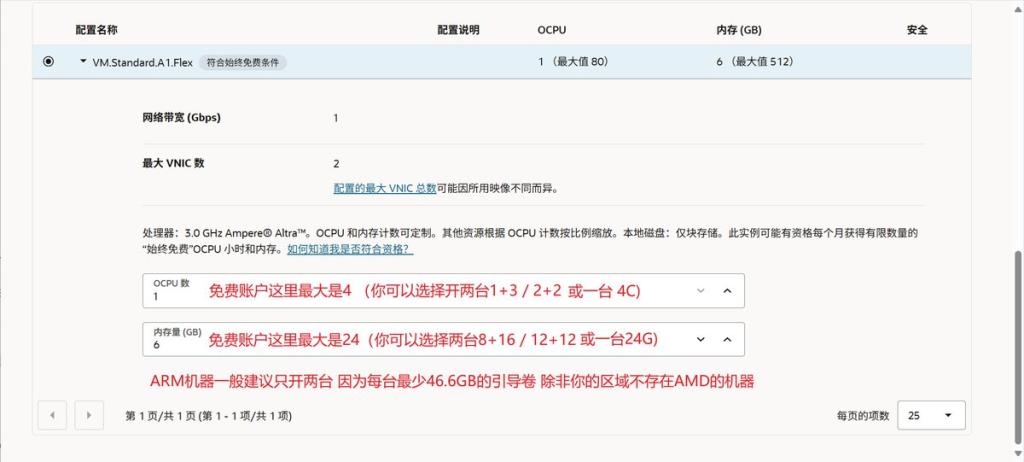
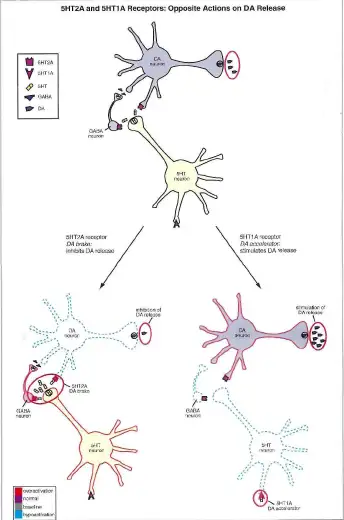
5-HT2A受体主要位于额叶/顶叶皮质和海马的谷氨酸神经元上,中脑的多巴胺能神经元上,垂体前叶的内分泌细胞上。
被激活后,它可以增加额叶/顶叶皮质中的谷氨酸能神经元,抑制多巴胺的释放,以及增加生长激素(GH) / 黄体生成素(ICSH ) / 促肾上腺皮质激素(ACTH)/ 催乳素(PRL)等的释放。
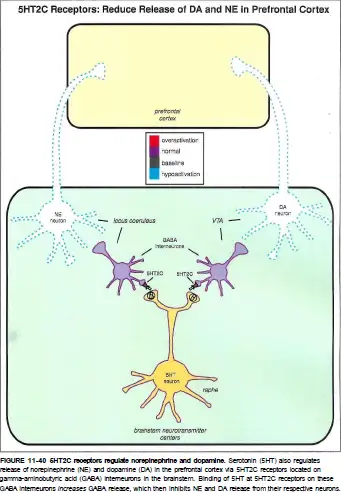
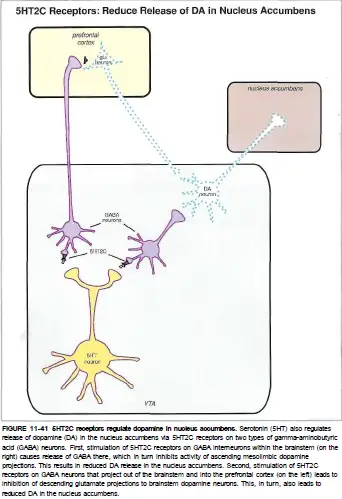
5-HT2C受体主要位于纹状体和下丘脑的多巴胺能神经元和去甲肾上腺素能神经元上。
被激活后,它可以抑制多巴胺和去甲肾上腺素的释放。
上述三种五羟色胺受体在情绪调节中发挥重要作用[1]。
具体的情况是复杂的,每个受体的功能都有两面性:
多巴胺和去甲肾上腺素的分泌被抑制,可以降低大脑皮层的活动并导致抑郁症状。 5-HT2C受体可以抑制多巴胺到伏隔核的释放,从而增加焦虑症状。
皮质中的谷氨酸能活性被抑制也会导致抑郁症状。
增加促肾上腺皮质激素(ACTH)的释放可以通过负反馈减少促肾上腺皮质激素释放激素(CRH)的释放,从而保护认知功能。因为促肾上腺皮质激素释放激素(CRH)可以导致海马神经元中Tau蛋白和轴突运输障碍的积累[2]。
此外,5-HT3受体和5-HT7受体也发挥着重要的作用:
5-HT3受体广泛位于中枢神经系统和消化系统,介导快速兴奋性突触传递和调节呕吐。
5-HT7受体的分布更广泛,它涉及温度调节,昼夜节律,记忆,睡眠,并在心血管系统和胃肠道内的平滑肌松弛中发挥作用。
值得注意的是,脊髓背角中的5-HT3受体和5-HT7受体在伤害性神经元的激活和抑制中起相反的作用[3] [4],5-HT3受体激动伤害性神经元而5-HT7受体抑制伤害性神经元。
5-HT4,5-HT5和5-HT6受体在神经系统调节中也有一些重要的作用。
由此,我们可以看到五羟色胺受体系统的特殊性。
在大多数情况下,两种相互拮抗的作用是由不同的神经递质介导的。例如,乙酰胆碱能交感神经和去甲肾上腺素能副交感神经的相互拮抗。
然而,许多拮抗作用都是由五羟色胺介导的,只不过涉及了不同类型的五羟色胺受体,这在其他神经递质中是非常罕见的。
比较不同神经递质受体的基因,研究发现:“原始”的五羟色胺受体在7.5亿年前已经开始进化[5]。除了5-HT3受体以外,大部分五羟色胺受体都是在那时开始分化。这个时间要早于毒蕈碱、多巴胺能和肾上腺素受体系统的进化。
这可能是基于五羟色胺本身的物理化学性质,使它更容易被细胞获得和合成,或者也可能完全是意外的因素导致的。总之,神经系统首先选择了五羟色胺受体来承担复杂的调节功能,由此形成了现在的五羟色胺受体系统。